In the beginning, there was no digestion, only ingestion. Simple, unicellular organisms just absorbed stuff through their outer membranes. Then along came multicellularity and, along with it, the need to increase the absorptive surface area for taking in nutrients. The simplest digestive systems around today consist of a simple digestive cavity, with one opening that serves as a mouth and an anus. Sounds gross to us, but that’s the way it is for organisms like hydra, jellyfish, planarians. They take in food items through their mouths, digest it inside a gastrovascular cavity, and then spit the undigestable bits back out of the mouth.
These simple, two-way digestive systems are often referred to as “incomplete” digestive systems — the idea being that complete digestive systems involve a one way flow of material from a mouth to an anus. As you can probably imagine, a one-way digestive system allows for a greater degree of efficiency. For instance, it allows an organism to eat while still digesting the previous meal. As a result, organisms with “complete” digestive systems can be more opportunistic in their feeding habits.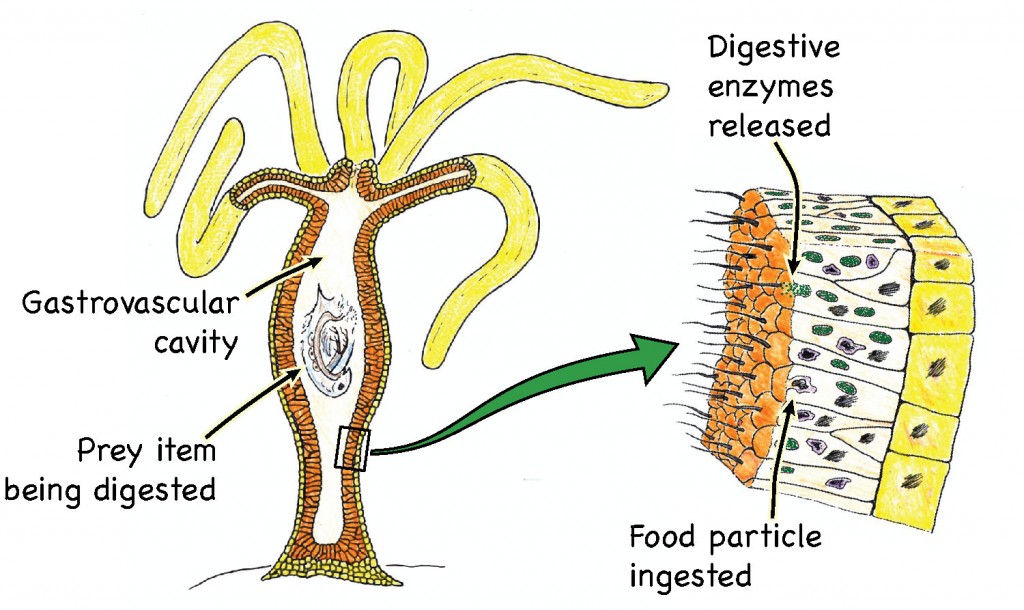
In general, digestive systems are highly adapted to coordinate with the feeding and foraging modes of the animal in question. For example, ruminants like cows, which feed on tough-to-digest plant material, have a particularly complex digestive systems involving symbiotic microbes that can break down cellulose from plant material. For ruminants, eating and digestion is a full time job, and their digestive systems are akin to glucose extraction factories. Other organisms have digestive systems geared toward a feast-or-famine cycle of food intake. For example, polar bears can eat up to 150 lbs at one sitting, and then fast for weeks with hardly a stomach rumble.
Clearly, digestive systems vary considerably across organisms. Among insects, the typical digestive system consists of the following:
- mouth,
- esophagus,
- crop
- stomach
- foregut
- hindgut
- anus

But there are some really interesting modifications to this general pattern. Blood-sucking insects like mosquitoes have to deal with a massive intake of salt and fluid. Their digestive systems will generally feature an enlarged storage vessel of some sort (in mosquitoes it’s the foregut) and adaptations for quickly removing excess fluid and salt. For instance, some mosquitoes begin excreting a salty liquid waste product before they have even finished biting their host1.

As organisms get bigger, they need to increase the absorptive surface of their digestive tracts. In most birds, mammals, and fish, increased surface area is accomplished by elongation of the intestines, which tend to become coiled into a complex knot of tubular tissue. However, in sharks and a few other elasmobranchs, we find what is called the spiral valve in the intestine. The spiral valve routes material through a sort-of spiral staircase in the intestine, which generates a lot more opportunity for absorption than a simple tube.

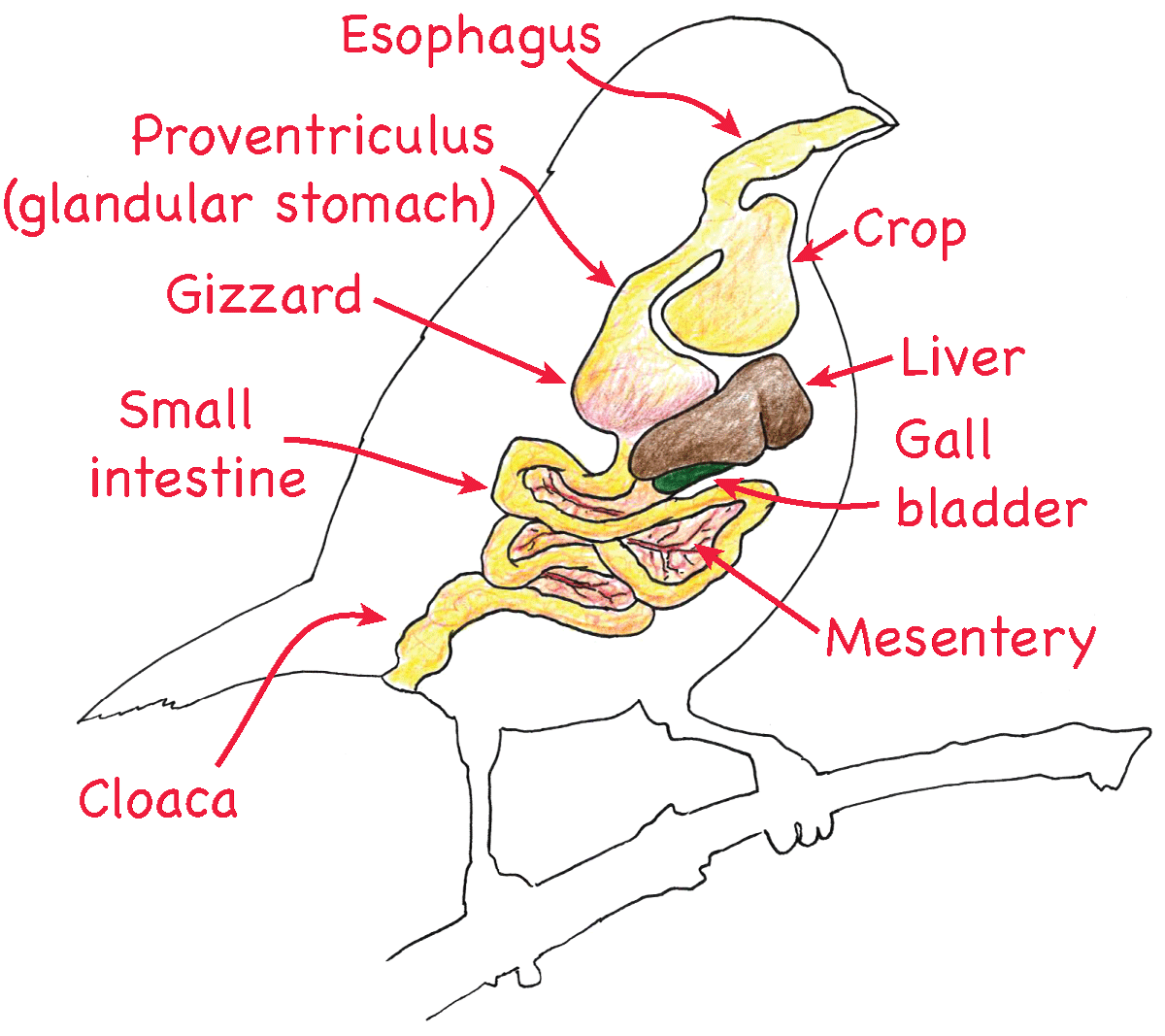
Many digestive systems feature a muscular stomach that breaks up food items both mechanically and chemically. Birds, since they lack teeth, rely heavily on their stomachs for mechanical digestion. Bird digestive systems are unusual due to the fact that they have no teeth. To compensate for their lack of dentition, birds typically have an extremely muscular and motile stomach or gizzard that performs mechanical digestion. Many birds will also swallow small stones that remain in the gizzard and help pulverize food.
Mammals
The process of digestion is, of course, most intensively studied in mammals. So let’s take the tour of the mammalian digestive system. Imagine that you are a bite of a peanut butter and jelly sandwich that is about to meet its intended destiny. The mouth opens, and in you go. You get coated with saliva and the teeth break you apart, which creates more surface area on which salivary enzymes can work. Amylases start to break apart all of your complex carbohydrates. This is your first experience of a reaction called hydrolysis.
Hydrolysis is a key process in digestion. Hydrolysis occurs when a water molecule combines with another molecule in such a way that both molecules get split up. The water molecule gives up one of its hydrogens, which replaces part of the other compound. The remaining OH atoms from the water molecule then bind to the part of the other molecule that was liberated by the hydrogen molecule.
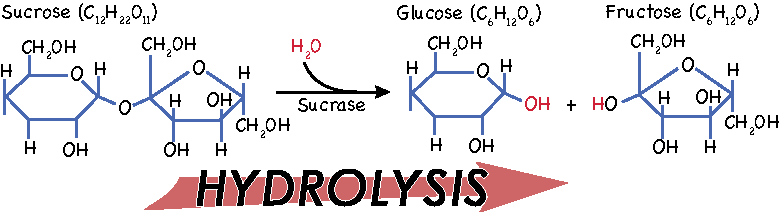
This process of splitting up molecules with water is how we separate relatively large molecules of carbohydrates, fats, and proteins into sugars, fatty acids, and amino acids that we can absorb. The process of hydrolysis is facilitated by a whole host of enzymes, but can also occur by manipulating acidity. We’ll see that digestive systems take advantage of both methods.
OK, so after some jostling and chewing, you have been packaged up into a mushy glob. The tongue pushes you toward the back of the throat, and this little flap of tissue — the epiglottis — folds down and suddenly you are in the esophagus and on your way to the stomach. It is not a fast, crazy plummet to the stomach. The smooth muscles surrounding the epiglottis contract in an orchestrated manner called peristalsis2, which gives you a smooth and controlled ride down.
Then it gets crazy. You get squeezed through the gastroesophageal sphincter and plunge into the body (or corpus) of the stomach. This is more or less the storage bin of the stomach. Farther down in the antrum is where the real action takes place. You barely have time to notice that the surface of the stomach is soft, slimy, and very wrinkly3 before the whole world (or what you can see of it) gets turned upside down. You realize that the muscles of the stomach wall are contracting and pushing you down farther into the stomach. The stomach starts to do it’s own version of peristalsis, but it’s a lot more intense that what happened in the esophagus. Meanwhile, fluid is being secreted from little holes in the stomach lining called gastric pits. There must be something in this fluid, because you begin to dissolve at an ever increasing rate. In fact, with all of this sloshing around in digestive enzymes on top of the chewing, you quickly cease to exist as a cohesive part of a PB&J sandwich. Therefore, for the rest of the tour, consider yourself to be the disembodied and omniscient spirit of a sandwich.
Using your newfound omniscience, you take a closer look at those gastric pits and it becomes apparent that three types of cells dominate the lining of the structure. There are mucous cells that secrete, well, mucous of course. And there are chief cells that secrete pepsinogen. What you ask yourself is pepsinogen? It is the precursor to a very active protein-digesting enzyme called pepsin. Pepsinogen gets transformed into pepsin when it comes into contact with an acid4. Then you notice that there is indeed a lot of acidity in the stomach, which you perceive to be the result of the third type of cell, parietal cells, pumping hydrogen and chloride ions out into the gastric pit5. Recalling your last chemistry class in sandwich school, you realize that these ions combine to make hydrochloric acid.

Looking even closer at these gastric pits, you see that there are a few other types of cells. One type of cell seems to be secreting histamine, which is normally associated with inflammatory responses. In this case, the histamine serves to stimulate the parietal cells to make more HCl. The cells secreting the histamines are called enterochromaffin-like cells (or just ECL cells). In other gastric pits, especially those toward the pyloric portion of the stomach (the far end), there are two other types of cells. One of these cell types, called G cells, is secreting gastrin, which is actually stimulating the activity of the parietal, chief, and ECL cells. Then there are the D cells. They don’t seem to be doing anything at the moment, but your intuition tells you that if the stomach works up too much acid, the D cells can secrete somatostatin (yes, the antagonist of growth hormone), which inhibits the activity of parietal, G, and ECL cells. In other words, D cells are the off switch.
So if the D cells are the off switch, where was the on switch? It was actually the central nervous system that got gastric secretion started, and it probably happened well before your descent into the digestive system. Simply seeing or smelling food can initiate an anticipatory response in the stomach that prepares it for a shipment of food. To be more specific, anticipation of eating leads to signals from the cerebral cortex and from the appetite centers of the amygdala and hypothalamus to stimulate the stomach via the gastric nerve. The signal reaches the cells of the stomach lining in the form of acetylcholine release from the enteric nervous system. Acetylcholine then initiates the activity of the chief cells, parietal cells, ELC cells, and G cells. This preparatory stage, which continues through all the chewing and swallowing that precedes the filling of the stomach, is known as the cephalic phase of gastric secretion. What follows is the gastric phase, and it centers around the stimuli that arise from actually having food in the digestive tract as opposed to stimuli from the external senses.
OK, so now the sandwich you once were has been reduced to a thick and acidic mixture we’ll call chyme. The stomach has done all it can do, so it’s time to pass everything on to the small intestine. The gatekeeper between the stomach and the intestines in the pyloric sphincter. This ring of smooth muscle is tonic — contracted to some extent all the time — and opens up only momentarily to allow the gradual passage of chyme into the upper portion of the small intestine (i.e., the duodenum).
The tissue of the small intestine is not equipped to deal with the acidity of the gastric secretions, so the duodenum has to have some means of neutralizing acid as the stomach contents empty into it. So the linings of the duodenum as well as the jejunum (the second segment of the small intestine) have S-cells that secrete the hormone secretin6. This hormone’s target is the pancreas, where it stimulates pancreatic duct cells to release sodium bicarbonate (NaHCO3) — yes, that’s the same thing as baking soda — which neutralizes the acid.
Secretin also has an inhibitory effect on gastric motility. That is, it slows the rate at which chyme enters the small intestine. This protective mechanism has to do with the osmolarity of chyme from the stomach. The stomach breaks down relatively large molecules into smaller ones, which dramatically increases the osmolarity of the chyme destined for the intestine. If it all got passed through to the duodenum at once, it would create an osmotic gradient that would draw a large amount of water in through the intestine and increase the volume of the fluid to the point that the intestine would become distended. So secretin helps put the brakes on the input from the stomach.
The small intestine must also be able to deal with fats coming from the stomach. Fats are hydrophobic and particularly hard to dissolve in aqueous solutions, and so the stomach does not do much to break down lipids. Once again, the duodenum has specialized sensory and secretory cells that can call in corrective measures. Cells in the duodenal mucosa called I-cells respond to high levels of fats (and to a lesser extent proteins) by secreting the hormone cholecystokinin (CCK). This hormone induces pancreatic acinar cells to release a cocktail of digestive enzymes for dealing with fats (i.e., lipases) and proteins (i.e., proteases).
Back when you were an intact sandwich, you had a fair amount of fat packaged into your layer of peanut butter. After some processing in the stomach, that fat is still present, mostly in the form of large fat droplets that have glommed together as a result of fat’s low solubility in water. The pancreatic lipases secreted in response to cholecystokinin make a little headway, but they need some help to break apart these large fat droplets. To fill this need, the digestive system mobilized a thin green fluid called bile. Bile is produced and recycled by the liver and gets stored in a small sac of tissue called the gallbladder. The gallbladder empties into the duodenum via the sphincter of Oddi, another ring of smooth muscle that regulates the timing of adding stuff to the small intestine.
A key ingredient in bile is bile salts. Bile salts act on the fat droplets similar to how detergents in dish soap act on the grease stuck to a frying pan. That is to say, bile salts are emulsifiers. Bile salts are derived from cholesterol, which gives them a hydrophobic (lipid soluble) side, and they also have charged, hydrophilic (water soluble) side derived from taurine or glycine. As the bile salts rumble around in the intestine with the large fat droplets, they begin to form shells around fat droplets with the hydrophobic end of the salts facing the fat and the hydrophilic ends facing outwards.
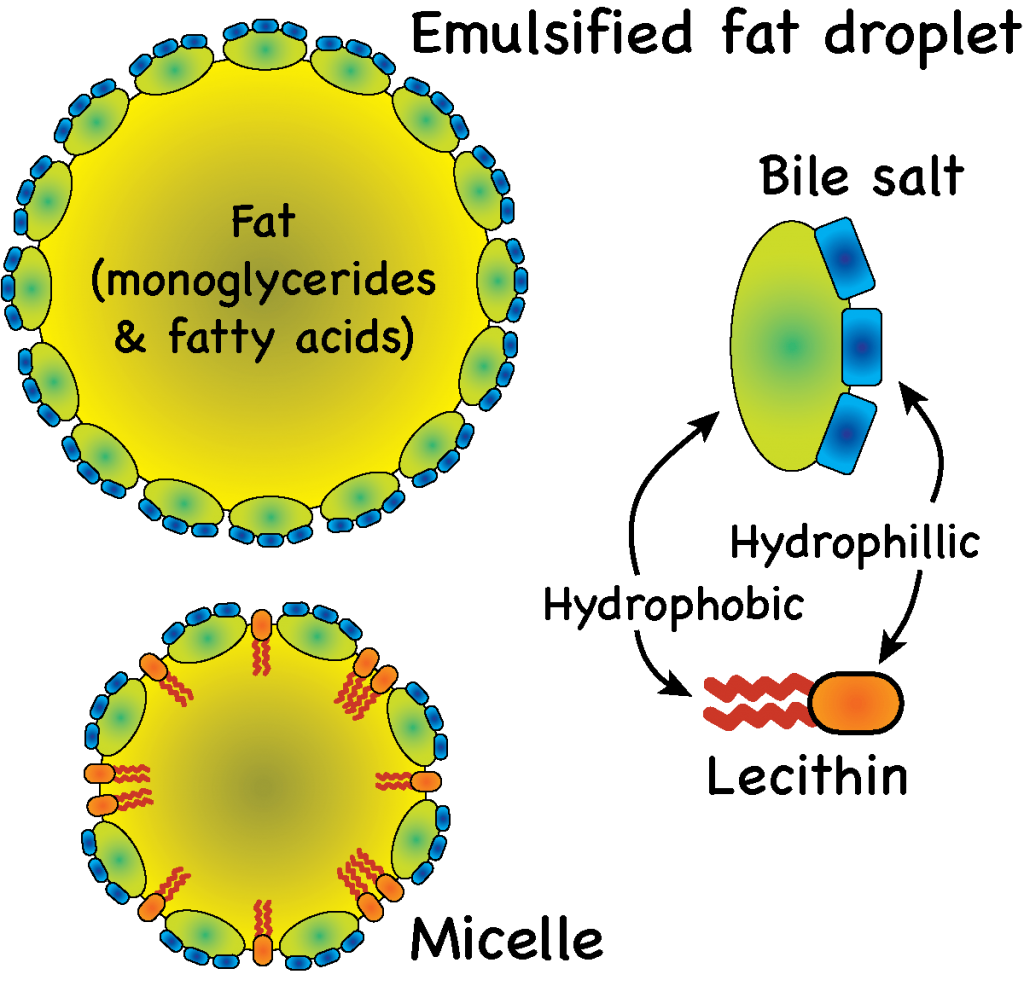
These emulsified droplet, called micelles, become increasingly small as mechanical (gut movement) and chemical (lipase) digestive efforts combine forces. At some point, lecithin, another component of bile, also joins the party. Lecithin is a phospholipid, with a polar head and a hydrophobic tail. Lecithin inserts itself into the micelles between the bile salts, helping to break up fat globules even further. As they get smaller, the surface area to volume ratio increases, allowing lipase to work very effectively in cleaving triglycerides into free fatty acids. Eventually they become small enough to get absorbed by the gut lining — more about that later.
An interesting thing about bile salts is that they get recycled. There are a host of specialized bile salt transporters 7 located throughout the lining of the small intestine (but mostly in the terminal portion (i.e., the terminal ileum). These transporters take up most of the free bile salts that are left behind after the fats they sequestered have been absorbed. About 5% of the bile salts are lost to the large intestine and eventually eliminated. But the other 95% are absorbed into the blood and then filtered out by the liver where they can go back into the digestive system again. In fact, a typical bile salt molecule will go through this process two or three times over the course of digesting a single meal.
OK, so what remains of the sandwich that once was you is now a soup full of peptides8, amino acids, short chains of carbohydrates, monosaccharides, emulsified fat droplets, and nutritive ions like Na+ and K+. What happens next is that all of this stuff has to somehow get absorbed by the cells of the intestinal epithelium and then passed along into the circulatory system. But before we get into that, let’s take a look at the lining of the small intestine. Apparent to the unaided eye are thousands of little finger-like extensions of tissue projecting out into the inside of the intestine (i.e. the lumen). Zooming in a little closer we see that each of these projections in covered in even smaller projections called villi (singular: villus). And then the cells that make up the luminal side of each villus have their own little microscopic projections called microvilli. This structure generates lots and lots of surface area for absorption. Thanks to all of these villi and microvilli, the total surface area of the small intestine is about the size of a tennis court.

For the sugars and amino acids, absorption occurs via secondary active transport, and the workhorse behind it all is the Na+/K+ ATPase pump. The basolateral membrane 9 is packed with Na+/K+ pumps, and they are constantly pumping Na+ out of the cell and into the interstitial fluid in the direction of the circulatory system. This makes for a relatively low concentration of Na+ inside the epithelial cells and generates a Na+ gradient with regard to the soupy stuff in the lumen of the intestine. The luminal membrane of the epithelial cells has several co-transport proteins that exploit this Na+ gradient to bring in other nutrients. For example, glucose and galactose are transported into epithelial cells via one of these co-transport proteins10. Most peptides and amino acids also pass into the intestinal epithelium via similar secondary active co-transport systems. There are a few exceptions here for some amino acids, which actually use an H+ gradient to get transported into the epithelial cells. What sets up the H+ gradient? Well, its actually secondary active antiport driven by that good ol’ Na+ gradient. So in the end, the Na+/K+ ATPase pumps in the basolateral membrane are doing all the work with regard to carbs, peptides, and amino acids. Each of these nutrient molecules coming into the cells generates its own concentration gradient with relatively high concentrations in the cell. So getting through the basolateral membrane is generally accomplished by simple facilitated diffusion through the appropriate transport protein. This passive movement of nutrients continues to carry these nutrients all the way into the capillaries that line the epithelial tissue.
Absorption of fats is a bit different. Recall that the fats have been packaged into micelles and broken down into monoglycerides and fatty acids. When the micelles come into contact with the epithelial cells, these molecules can simply pass through the luminal membrane via passive diffusion. Once inside the epithelial cells, the fat components are reassembled into triglycerides. These triglycerides are then aggregated together into tiny droplets that are then passed through the endoplasmic reticulum, which creates a thin shell of lipoproteins around the glob of triglycerides. The lipoprotein coat makes the glob water soluble, and it is actually called a chylomicron11 at this point, not a glob. The chylomicrons are actively transported through the basolateral membrane via exocytosis. They are too big to get into the capillaries, and so they get taken up by the lymphatic system. From there, they may be dumped into the circulatory system and conducted to hardworking cells to be used as an energy source.
After the small intestine is through with the nutrients in that PB&J, there’s not much left. But we are not done yet. The next stage has what remains of your sandwich remains passing through the ileocecal valve into the cecum of the large intestine. Here, the pace slows down a bit. With the large intestine or colon, there no great rush to obtain as many nutrients as possible as quickly as possible. Hence, there’s a lot less motility. Brief haustral contractions12 occur from time to time to mix things up a bit, and there are occasional waves of peristalsis to move things along. Because of this slow pace of activity, the large intestine can host activities like fermentation to release more useful material from that PB&J. Fermentation involves bacteria, and most digestive systems have their own unique sets of bacteria or gut flora that help release nutrients, vitamins, and minerals that were out of reach for the small intestine. In many cases, these bacteria actually synthesize special fatty acids13 or vitamins14. There are really no digestive enzymes released in the mucous solution with alkaline buffers that make the colon a nice place for bacteria.
Hours pass and all that’s left of your physical form is a little bit of indigestible cellulose. It is now mixed with bilirubin and biliverdin15, a little salt, some water and bacteria. Together these are the ingredient of feces, and they are ready for defecation. But nothing’s happening. It’s all just sitting here.
Then it happens. Suddenly the large intestine comes to life and all of the waste material gets rapidly pushed toward the lower end of the large intestine. What has happened is that the person who ate you has just made herself another sandwich. Eating can often cause a gastrocolic reflex in response to gastrin secretion by the stomach. This reflex involves strong peristaltic waves in the large intestine that move its contents into the rectum, which is the final stop before defecation.
This filling of the rectum stimulates stretch receptors that cause the internal anal sphincter to relax. They also send a message to the brain that it is time to find a bathroom or other appropriate place. The only thing preventing defecation at this point is the external anal sphincter, which is a skeletal muscle that is under voluntary control. When this muscle is relaxed, the contents of the rectum are emptied. This is usually accompanied by contractions of the abdominal muscles and a forced expiration of the lungs against a closed glottis, both of which generate pressure in the lower abdomen.
For more details on the anatomy and basic functions of the digestive system components please refer to the following sections of Openstax Anatomy and Physiology:
The three stages of digestion:
Hormonal control of gastric secretion:
- Brief overview of digestive hormones and regulation from the Sciencelearn Hub